AWA: Academic Writing at Auckland
Title: Proposal to develop transgenic species for biofuel production
|
Copyright: Stephanie Morton
|
Description: Write a research proposal for the development of a transgenic plant that could be used to enhance biofuel production.
Warning: This paper cannot be copied and used in your own assignment; this is plagiarism. Copied sections will be identified by Turnitin and penalties will apply. Please refer to the University's Academic Integrity resource and policies on Academic Integrity and Copyright.
Proposal to develop transgenic species for biofuel production
|
Title of the Project: Lignin reduction in maize for enhanced biofuel production Summary: Lignocellulose is one of the most abundant sources of energy available on the planet but there is a significant difficulty with accessing this resource in a cost efficient way that also achieves a net energy gain through biofuel processing. Second generation biofuels, which utilise the sugars and starches locked in the cell walls of plants by lignin, cellulose, and hemicellulose structures, require physical and chemical pre-treatments to be broken down before biofuel fermentation can begin. Reductions in these cell wall components have shown to increase digestibility of biomass and increase relative biofuel energy output. Caffeic acid O-methyltransferase (COMT) and Caffeoyl-CoA-3-0-methyltransferase (CCoAOMT) facilitate guaiacyl (G) and syringyl (S) lignin biosynthesis within the cell wall of maize (Zea mays). This study takes a transgenic approach to reduce the total and relative S and G lignin within the cell wall. COMT and CCoAOMT gene promoters will be targeted with an Agrobacterium mediated gene transfer of CRISPR-Cas9 complex. Each enzyme gene promoter will be targeted in separate maize plants to study the effects of S/G lignin ratio reduction via COMT promoter knockout, and combined S and G lignin reduction via CCoAOMT promoter knockout.
BACKGROUND
Biofuels are a renewable form of energy derived from the fermentation of biomass into ethanol and biodiesel (Highina et al., 2014; Wang et al., 2016, Renzaho et al., 2017). With global energy needs rising and CO2 emissions of major concern biofuels are seen as a clean and sustainable solution to the growing problem (Renzaho et al., 2017). First generation biofuels utilise the sugars, starches, and plant oils primarily from the edible parts of arable crops, and are contributing significantly to global energy needs already (Wang et al., 2016). Second generation biofuels aim to utilise non-edible parts of the plant by unlocking the lignocellulose inside the plant cell wall (Mansfield, 2009; Wang et al., 2016). Lignocellulose is the most abundant source of convertible biomass available producing 1.5 x1010 tons/year (Wang et al., 2016). However biological degradation of lignocellulose is not as efficient at solubilizing the carbohydrates away from other cell wall constituents. The biomass must first go through physical and chemical pre-treatments in order to break down the cell walls and release the lignocellulose, which makes the process expensive with only a 60% energy conversion rate (Mansfield, 2009; Wang et al., 2016). The pre-treatments such as dilute acid, steam explosion, AFEX or organosolv are typically required to assist the carbohydrate release (Mansfield, 2009).
Primary plants used for second generation biofuels are grasses such as switchgrass (Panicum virgatum L.), but there is a need to look into more efficient sources such as maize stover (Baxter et al., 2016). Maize stover is widely accessible across Europe and USA for forage, each year around 8.6 MHa is grown (Chen et al., 2014). Maize cob is currently used for first generation biofuels but it is inefficient with a 29% net energy loss in conversion (Pimental, 2003). Maize cob biofuels also compete with food production and create an inefficient use of land coverage (Wang et al., 2016; Renzaho et al., 2017). So research is looking at ways of utilising the maize stover for biofuels whilst keeping the cob for food production. Lignocellulose is available in the cell wall of the stover. The three primary structural units that create challenges for unlocking the lignocellulose for biofuel production are cellulose, hemicellulose, and lignin (Mansfield, 2009). Lignin’s are the focus of this project as they show promising results in increasing cell digestibility.
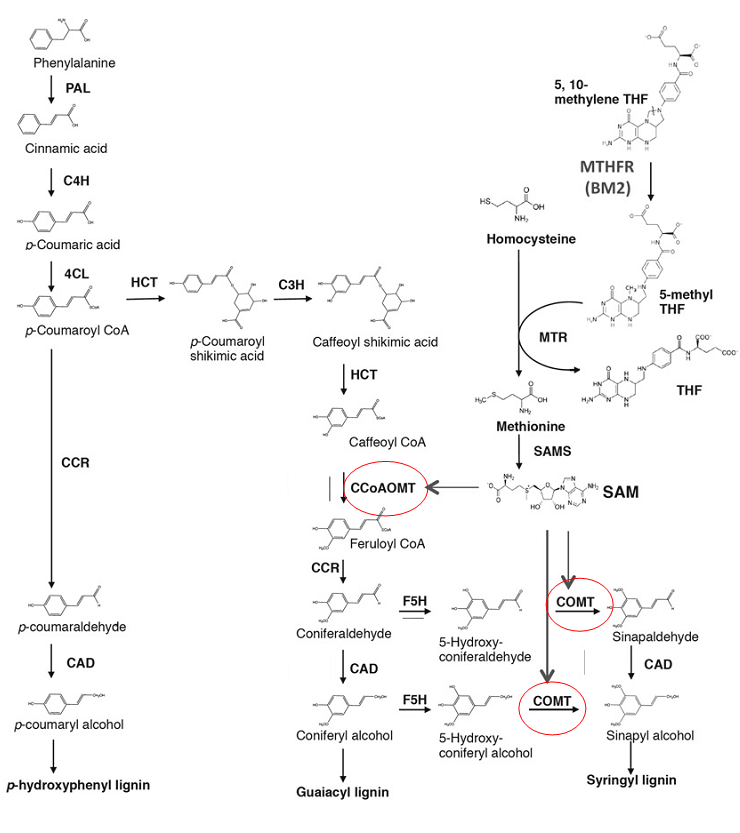
Lignin is an important structural component of the plant secondary cell wall that cross-links cell wall polysaccharides (Boerjan et al., 2013; Mansfield, 2009; Oraby and Ramadan, 2014). Lignin’s are complex racemic aromatic heteroploymers that have an important function in structural support, defence and water retention of the cell which help resistance to degradation (Capellades et al., 1996; Boerjan et al., 2013; Beckham et al., 2016). The lignin biosynthesis pathway starts after the Shikimate pathway where the deamination of L-phenylalanine creates cinnamic acid (see figure 1). Successive steps create three hydroxycinnamyl alcohol monomers that differ in degree of methoxylation. These lead on to the production of three monomeric subunits of lignin; p-hydroxyphenyl (H), guaiacyl (G), and syringyl (S) (Li et al., 2013). Lignin makes up 13-21% of the cell wall polymer composition (Wang et al., 2016). Guaiacyl and syringyl lignin account for >90% of this lignin in angiosperms and only differ in chemical structure by one or two methoxyl groups (Fornale et al., 2017).
There are 10 enzymes required in the biosynthesis of monlignols (Li et al., 2013) and manipulation of these to create a more digestible plant have been a focus in recent years (Wang et al., 2016), in particular the Cinnamyl alcohol dehydrogenase (CAD), Caffeic acid O-methyltransferase (COMT) and Caffeoyl-CoA-3-0-methyltransferase (CCoAOMT) which play different roles in regulating H,G and S lignin (Sattler and Funnell-Harris, 2013). Research has looked into Lignin synthesis, interlinking, and regulation across a few different species including alfalfa, Arabidopsis, poplar, rice, sorghum, sugar cane, and switchgrass (Wang et al., 2016). The evidence suggests that reduction of lignin content in
Figure 1. The lignin biosynthesis pathway displaying the location of COMT and CCoAOMT in guaiacyl (G) and syringyl (S) lignin synthesis (Tang et al., 2014).
the cell wall is able to increase digestibility but with mixed results of yield and structural stability of the plant (Wang et al., 2016). Much has been learnt from existing mutants in maize (brown midrib 1,2,3, and 4 mutants) as to the lignin biosynthesis pathway and the effects CAD, COMT and CCoAOMT have on regulating H, G and S lignin (Piquemal et al., 2002; Sattler and Funnell-Harris, 2013). Low levels of lignin reduction achieved through RNAi experiments on the COMT gene, mimicking the maize natural mutants, have produced similar results with increased digestibility as well as producing reddish brown pigment in lignified tissue, while also reducing S/G ratios (Piquemal et al., 2002). Other experiments with RNAi on CCoAOMT have produced 10.35% reductions in Klason lignin and increases in S/G ratios (Li et al., 2013).
The aim of this project is to look for ways of reducing, through genetic modification with the CRISPR Cas9 system, the expression of COMT and CCoAOMT in maize with the intention of designing a more easily degradable and cost efficient crop to process into biofuels (Mansfield, 2009; Wang et al., 2016).
PROPOSED PROJECT
The strategy of this project will be to elicit individual knock outs of the COMT and the CCoAOMT genes in maize (Zea mays) with the CRISPR-Cas9 system. The purpose is to reduce the level of S-lignin and S&G lignin respectively within the cell wall thus making cells easier to break down via physical and chemical pre-treatments used in the biofuel processing of lignocellulosic biomass (Beckham et al., 2016). Maize contains a single COMT gene coding for one enzyme in the lignin biosynthetic pathway described by Collazo and colleagues (1992) and two closely related homologs (93.1%similar at the amino acid level) of the CCoAOMT gene (GRMZM2G099363 and GRMZM2G127948) (Wang and Balint-Kurti, 2016). This study will involve the transformation of the maize DNA using Agrobacterium tumefaciens. We will insert a Cas9 protein into the nucleus with guide RNA sequences specifically targeted to the promoter regions of the CCoAOMT and COMT genes (Collazo et al., 1992; Zhu et al., 2016; Addgene, 2017). In Zea mays Caffeic acid O-methyltransferase (COMT) methylates 5-hydroxy-coniferaldehyde into sinapaldehyde and leads on to syringyl alcohol and S lignin production. Reduction in COMT results in reduction in S lignin only, completely deficient COMT maize have severely depleted S lignin but not absent suggesting there is supplementary pathway for S lignin (Barrie`re et al. 2004). Reductions of 8-12% lignin in the maize cell walls still maintain phenotypic structural rigidity, higher levels produce weakened cell walls (Barrie`re et al. 2004). Caffeoyl-CoA-3-0-methyltransferase (CCoAOMT) is an integral part of lignin biosynthesis. It is present earlier in the pathway catalysing the methylation of caffeoyl CoA to feruloyl CoA which in successive steps produces two monolignols; coniferyl and syringyl alcohols which lead on to production of S and G lignin’s (Barrie`re et al. 2004).
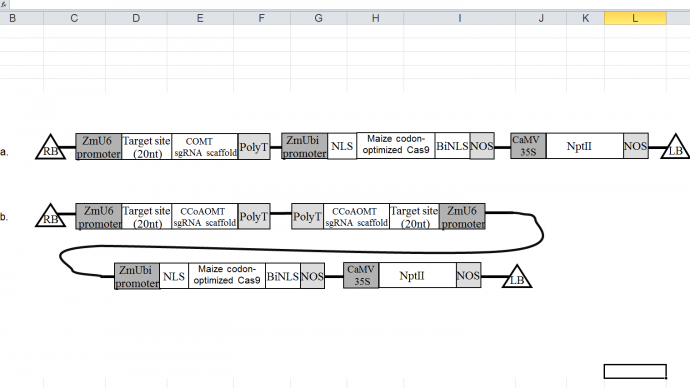
Construct overview Two constructs will be designed; one with a COMT promoter targeted sgRNA attached to Cas9 and selectable marker, the second with two CCoAOMT promoter targeted sgRNA attached to Cas9 and selectable marker (see figure 2). Constructs will be transferred into separate Zea mays plants as DNA vectors in one Agrobacterium transfer event (Svitashev et al., 2015, Char et al., 2017). The CRISPR-Cas9 system, once inserted into the nucleus, will transcribe a Cas9 nuclease protein which will use the associated sgRNA to locate the promoter regions of the CCoAOMT and COMT gene and cut the DNA eliciting a promoter knock out via non homologous end joining (NHEJ) (Zhu et al., 2016; Addgene, 2017).
Figure 2. a. Agrobacterium mediated vector construct containing a 20nt target site with sgRNA sequence for the COMT promoter, ZmU6 promoter and PolyT terminator, A maize codon optimized Cas9 with ZmUbi promoter, two nuclear localisation signals, and Nopaline synthase (NOS) terminator. Neomycin phosphototransferase II (NPT II) selectable marker with CaMV 35S promoter, Nopaline synthase (NOS) terminator, and left/right boarders .
The promoters- Three proven promoters in maize will be used for optimum transcription; ZmUbi (maize Ubiquitin2), ZmU6 (maize RNA polymerase III), and CaMV 35S (cauliflower mosaic virus promoter) ( Zhu et al., 2016). A maize Ubiquitin2 promoter will be used to initiate the Cas9 reading frame, and a maize ZmU6 snRNA polymerase III promoter will be used to initiate the sgRNA as previously described in Zhu and colleagues (2016). The CaMV 35S promoter will drive expression of the NPTII gene. CaMV 35S is a strong constitutive overexpression promoter that is commonly used in transgenic plants. It has been chosen because it has been shown to drive transgene expression in all parts of the maize plant, and raises the enzyme activity of NPTII and tolerance of kanamycin without creating instability in the expression of NPTII (Huang and Dennis, 1989; Miki and McHugh, 2004; Zahradnik et al., 2014).
Nuclear localisation facilitation- The gene coding Cas9 will have a nuclear localisation signal (monopartite SV40) attached to the N terminus, and a bipartite nucleoplasmin nuclear localisation signal added to the C terminus to facilitate nuclear localisation (Zhu et al., 2016). Two NLS’s are used in this construct to maximise delivery into the nuclei (Imagawa et al., 2000; Belhaj et al., 2013)
Terminator sequence- The terminator sequence of the Nopaline synthase (Nos-term) gene of A. tumefaciens will be used for termination of the Cas9 sequence and the selectable marker sequence. Nos-term is a common terminator sequence that has been successfully used in previous studies of transgenic maize (Huang and Dennis, 1989, Zhu et al., 2016). A PolyT terminator will be used for the guide RNA sequence as described in Zhu and colleagues (2016).
Selectable marker- Neomycin phosphototransferase II (NPT II) is a stable marker gene widely used in transgenic studies and highly capable of successfully integration into Maize (Huang and Dennis, 1989; Miki and McHugh, 2004). NPT II expression creates Kanamycin resistance within the transgenic plant (Miki and McHugh, 2004) that will be selected for by growing plants on a Kanamycin containing media to filter out non-mutants after transformation. NPT II has been chosen because is has no adverse effects on human health and is approved by the US Food and Drug Administration (FDA) (Miki and McHugh, 2004).
Guide RNA- Part of the experimental design will be to determine three sgRNA sequences, one for targeting the COMT promoter and two for targeting the homologues of CCoAOMT. This will be achieved following the protocols in CRISPR 101: A Desktop Resource (2nd Edition) manual (2017). Step1: Locating cDNA with desired expression pattern with the oligocapping method and genome sequencing of the COMT and CCoAOMT gene regions via microarray. Step 2: Selection of COMT and two CCoAOMT promoter sequences and cloning by library screening and checking for expression by GUS/GFP fusion. Step 3: Identification of most efficient ‘on target’ PAM sequence cleavage site within target gene regions (see cas9 binding site section). Step 4: Synthesis via PCR to generate template DNA and cloning desired sgRNA into the A. tumefaciens vector.
Target sequence-Protospacer sequence 20nt long will be inserted at the 5’ end of the gRNA. When locating the target sequence high GC content (40-80%) will be prioritised to stabilize the RNA:DNA duplex and destabilize off target hybridization. If this is not available TATA boxes or CAAT boxes will be targeted.
Transient assay- The activity of the CRISPR-Cas9 system will be tested in transient prior to Agrobacterium transformation to asses’ expression activity. Agrobacterium will be used to insert a COMT promoter fused to a Luciferase marker to asses expression patterns across the leaves and stem of tobacco plants (Nicotiana tabacum var. Xanthi) (Barnes, 1990; Velez-Bermudez et al., 2015). Tobacco plants will be grown in lab on growth media until 15-40cm tall, whole young leaves from nodes 3-6 down from the apex will be analysed under luciferase photoassay (Barnes, 1990). A standard CaMV 35S promoter will be used to regulate transcription. If this method does not produce viable results then rice protoplast will be used (Miao et al., 2013).
Cas9 variant- the Streptococcus pyogenes Cas9 (spCas9) variant will be selected to elicit a gene knockout via non-homologous end joining (NHEJ). SpCas9 is dependent on the 5’NGG 3’ Protospacer adjacent motif (PAM) sequence. If a 5’NGG 3’ PAM cannot be located a synthetic Cas9 variant with novel PAM recognition will be used to match the maize gene sequence (Addgene, 2017).
Cas9 Binding site- Protospacer adjacent motif (PAM) binding nucleotides will flank the 3’ end of the crRNA-targeted sequence (Svitashev et al., 2015; Ceasar et al., 2016). PAM nucleotides (5’NGG3’ for SpCas9) will be derived from unique existing sequences found in the genome (Zhu et al., 2016; Addgene, 2017). If this cannot be located, another Cas9 variant will be selected. This will not be determined until further genome sequencing is undertaken.
Transfer method- Agrobacterium tumefaciens binary expression vector will be used to transfer the construct into maize immature embryos. Methods will follow the techniques of Davey, Rech and Mulligan (1989) with adaptations to the CRISPER-Cas9 system (Addgene, 2017; Char et al., 2017). In the COMT T-DNA, the target site and sgRNA sequence with their own promoter and terminator will be inserted into the vector along with the gene of interest (Cas9) and selectable marker. In the case of CCoAOMT T-DNA for both target sites and sgRNA combinations will be simultaneously inserted into one Agrobacterium containing a single Cas9 gene with individual ZmU6 promoters and PolyT terminators (Char et al., 2017). Agrobacterium DNA vectors were chosen over RNP complexes delivered by biolistic-transformation due to greater stability and accuracy, higher mutation frequency, and lower cost of equipment (Svitashev et al., 2015; Svitashev et al., 2016; Char et al., 2017).
Generating the transgenic crop- Agrobacterium vector transformation and targeted CRISPR based mutagenesis will follow similar methods and estimated time frames set out by Char and colleagues (2017): ->Design of Cas9/sgRNA and Agrobacterium vector. (~2 weeks) ->Transformation of immature maize embryos via Agrobacterium vector. (~8 weeks) ->Screening for mutated callus lines by growing callus’s on kanamycin containing media. (~1 week) ->Regeneration of mutated callus lines into multiple plantlets. Based on the work of Zhu and colleagues (2016) it is estimated that approximately 10% of To Cas9 transformed plants will have NHEJ mutations in the COMT promoter, however this could range from 2%-100% (Char et al., 2017). We will run 500 lines for each target gene COMT and CCoAOMT. (~4 weeks) ->Identification of mutated plantlets with high throughput SNP genotyping of maize using the GoldenGate assay (Yan et al., 2010; Vad-Nielsen et al., 2016), it is anticipated 20-60 lines will contain the desired gene insertion whilst maintaining structural rigidity of the cell for further analysis. (~1 week ->Seed propagation via crossing and progeny analysis. (~12 week) ->Desired lines will be selected and the promoter regions undergo PCR amplification using primer pairs flanking the designated target sites (Zhu et al., 2016) and Sanger sequencing to genotype the promoter. (~1 week)
Glasshouse trials- 8 plants from To lines will be chosen for glasshouse trials based on percentage of lignin reduction (WT, ~10%, ~20%, and 30-40%). Backcrossing with wild type maize will be done to achieve optimum stable traits for analysis, and select out the Cas9/sgRNA/NPTII transgene (Pichon et al., 2006; Char et al., 2017). Southern blots under standard conditions will determine genetic status before moving to field trials. (~1 year)
Field testing- Field tests will be undertaken in the USA beginning April. Eight plots (one per progeny) will be planted in 6m long double rows with 0.75m spacing and 90,000 plants/Ha densities. A 15 m wide fallow and buffer barrier of unrelated species (e,g. soy) will surround the plot. Plants will be harvested at end of growing season (October-November) by hand and separated into genotype groups. Four plants from each plot (total 32 individual plants) will be sampled at mid growing season and again at harvest (Baxter et al., 2014). Agronomic performance will be measured by recording height and diameter at mid and end season, and dry weight biomass yield at end of season (Baxter et al., 2014). Chemical analysis will be run on individual plant stover (stalk and leaves) testing for lignin content and composition and averaged (Pichon et al., 2006). Cell wall residues will be analysed with Pyrolysis molecular beam mass spectrometry (py-MBMS) for total lignin and S/G monomer ratio for whole above ground biomass harvested at end of growing season (Baxter et al., 2014). Sugar release efficiency will be measured via enzymatic hydrolysis, and ethanol yield via simultaneous saccharification and fermentation (SSF) approach. (~1 year)
RISK ANALYSISStrength and yield- The most notable risk is the effects on cell wall rigidity and the ability of the plants to retain structure and maintain yields. As lignin is integral to the cell wall reductions and ratio changes in H, G, and S lignin could cause growth issues. However, previous studies in switchgrass (Panicum virgatum L.), poplar (Populus spp.), Brassica napus, Sorghum, and existing maize COMT mutants (bm2,3) have shown that a knock out/down of COMT leads to lower lignin content, increased biofuel digestibility, and little or no effect on yield (Vignols et al., 1995; Piquemal et al., 2002; Barrière et al., 2004; Sattler et al., 2012; Rubinelli et al., 2013; Oraby and Ramadan, 2014; Tang et al., 2014; Baxter et al., 2016). Further experiments in switchgrass have shown successive generations stably maintaining 11-16% decrease in lignin with lower S/G lignin ratios (33-40%) and 15-42% higher sugar release (Baxter et al., 2016). In general lignin levels were found to fluctuate over time and plants with 8-12% lignin down regulation grew normally, plants with 40% less lignin did not survive the first winter in the field (Pichon et al., 2006; Baxter et al., 2016). We will be focusing on a range of COMT and CCoAOMT expression levels for this reason. Some studies have also suggested that ratio changes in G and S lignin are counteractive to digestibility due to increases in coniferaldehyde units inhibiting digestibility up to 12% (Barrie`re et al. 2004; Fornale et al., 2017). Complete knockdown of COMT causes severe ratio changes, but not complete elimination of S lignin as the bm3 maize mutant still displays some incorporation of S-lignin monomers suggesting partial compensation of ZmCOMT by other methyltransferases (Fornale et al., 2017). For this reason we are testing CCoAOMT knockout as well as some studies show it has even effects on decreasing S and G lignin and may be better alternative (Barrie`re et al. 2004). Other studies in maize with RNAi downregulation also suggest CCoAOMT reduces Klason lignin content and significantly increased S/G ratio whilst maintaining normal phenotypes compared to wild types (Li et al., 2013). Testing both genes should provide more phenotypic variety of desirable traits to select from.
Plant health- Lignin reduction could also affect plant health as it is involved in immunity to biotic stress and wounding (Velez-Bermudez et al., 2015, Wang and Balint-kurti, 2016). In maize a hypersensitive response (HR) is induced by pathogen attack which increases expression of almost all genes in the lignin pathway in order to confine pathogens and prevent spread to other cells (Sattler and Funnell-Harris, 2013; Wang and Balint-kurti, 2016). But cases show that suppression of COMT and CCoAOMT have little/no and sometimes increased resistance to biotic stress, and H lignin is what tends to elevate and be required for defence (Sattler and Funnell-Harris, 2013; Miedes et al., 2014; Baxter et al., 2016). Therefore we anticipate a low risk to disease immunity in the genetically modified maize. Cross pollination and human consumption- As this experiment involves a gene knockout and Cas9/sgRNA/selectable marker will be crossed out before field trials the probability of cross pollination or horizontal gene transfer of the construct to other crops is unlikely. Spread of mutations in COMT and CCoAOMT gene promoters are also of minimal concern as lignin reductuced mutants already exist in maize and appear to be disadvantageous traits in wild types (Barrie`re et al. 2004; Vignols et al., 1995; Piquemal et al., 2002). There is little risk to human consumption as the cob will not be affected. Lignin is not produced in the cob and the Cas9/sgRNA/selectable marker construct will be crossed out.
REFERENCES
Addgene. (2017). CRISPR 101: A Desktop Resource (Vol. 2nd Edition, pp. 1-195): Addgene. Barnes, W. M. (1990). Variable Patterns of Expression of Luciferase in Transgenic Tobacco Leaves. Proceedings of the National Academy of Sciences of the United States of America, 87(23), 9183-9187.
Barrière, Y., Ralph, J., Méchin, V., Guillaumie, S., Grabber, J. H., Argillier, O., . . . Lapierre, C. (2004). Genetic and molecular basis of grass cell wall biosynthesis and degradability. II. Lessons from brown-midrib mutants. Comptes Rendus Biologies, 327(9-10), 847-860. doi:10.1016/j.crvi.2004.05.010
Baxter, H. L., Mazarei, M., Fu, C., Cheng, Q., Turner, G. B., Sykes, R. W., . . . Stewart, C. N. (2016). Time Course Field Analysis of COMT-Downregulated Switchgrass: Lignification, Recalcitrance, and Rust Susceptibility. BioEnergy Research, 9(4), 1087-1100. doi:10.1007/s12155-016-9751-1
Beckham, G. T., Johnson, C. W., Karp, E. M., Salvachua, D., & Vardon, D. R. (2016). Opportunities and challenges in biological lignin valorization. Curr Opin Biotechnol, 42, 40-53. doi:10.1016/j.copbio.2016.02.030
Belhaj, K., Chaparro-Garcia, A., Kamoun, S., & Nekrasov, V. (2013). Plant genome editing made easy: targeted mutagenesis in model and crop plants using the CRISPR/Cas system. Plant Methods, 9, 39. http://doi.org.ezproxy.auckland.ac.nz/10.1186/1746-4811-9-39
Boerjan, W., Ralph, J., & Baucher, M. (2003). Lignin biosynthesis. Annu Rev Plant Biol, 54, 519-546. doi:10.1146/annurev.arplant.54.031902.134938
Capellades, M., Torres, M. A., Bastisch, I., Stiefel, V., Vignols, F., Bruce, W. B., . . . Rigau, J. (1996). The maize caffeic acid O-methyltransferase gene promoter is active in transgenic tobacco and maize plant tissues. Plant Molecular Biology, 31(2), 307-322. doi:10.1007/bf00021792
Ceasar, S. A., Rajan, V., Prykhozhij, S. V., Berman, J. N., & Ignacimuthu, S. (2016). Insert, remove or replace: A highly advanced genome editing system using CRISPR/Cas9. Biochim Biophys Acta, 1863(9), 2333-2344. doi:10.1016/j.bbamcr.2016.06.009
Char, S. N., Neelakandan, A. K., Nahampun, H., Frame, B., Main, M., Spalding, M. H., . . . Yang, B. (2017). An Agrobacterium-delivered CRISPR/Cas9 system for high-frequency targeted mutagenesis in maize. Plant Biotechnol J, 15(2), 257-268. doi:10.1111/pbi.12611
Chen, Y., Blanco, M., Ji, Q., Frei, U. K., & Lubberstedt, T. (2014). Extensive genetic diversity and low linkage disequilibrium within the COMT locus in maize exotic populations. Plant Sci, 221-222, 69-80. doi:10.1016/j.plantsci.2014.02.004
Collazo, P., Montoliu, L., Puigdomènech, P., & Rigau, J. (1992). Structure and expression of the lignin O-methyltransferase gene from Zea mays L. Plant Molecular Biology, 20(5), 857-867. doi:10.1007/bf00027157 Davey, M. R., Rech, E. L., & Mulligan, B. J. (1989). Direct DNA transfer to plant cells. Plant Molecular Biology, 13(3), 273-285. doi:10.1007/bf00025315
Fornale, S., Rencoret, J., Garcia-Calvo, L., Encina, A., Rigau, J., Gutierrez, A., . . . Caparros-Ruiz, D. (2017). Changes in Cell Wall Polymers and Degradability in Maize Mutants Lacking 3'- and 5'-O-Methyltransferases Involved in Lignin Biosynthesis. Plant Cell Physiol, 58(2), 240-255. doi:10.1093/pcp/pcw198
Highina, B. K., Bugaje, I. M., & Umar, B. (2014). A review on second generation biofuel: a comparison of its carbon footprints. European Journal of Engineering and Technology Vol, 2(2).
Huang, Y.-W., & Dennis, E. S. (1989). Factors influencing stable transformation of maize protoplasts by electroporation. Plant Cell, Tissue and Organ Culture, 18(3), 281-296. doi:10.1007/bf00043398
Imagawa, M., Sakaue, R., Tanabe, A., Osada, S., & Nishihara, T. (2000). Two nuclear localization signals are required for nuclear translocation of nuclear factor 1-A. FEBS Letters, 484(2), 118-124. doi:10.1016/S0014-5793(00)02119-0
Li, X., Chen, W., Zhao, Y., Xiang, Y., Jiang, H., Zhu, S., & Cheng, B. (2013). Downregulation of caffeoyl-CoA O-methyltransferase (CCoAOMT) by RNA interference leads to reduced lignin production in maize straw. Genetics and Molecular Biology, 36(4), 540–546. http://doi.org.ezproxy.auckland.ac.nz/10.1590/S1415-47572013005000039
Mansfield, S. D. (2009). Solutions for dissolution--engineering cell walls for deconstruction. Curr Opin Biotechnol, 20(3), 286-294. doi:10.1016/j.copbio.2009.05.001
Miao, J., Guo, D., Zhang, J., Huang, Q., Qin, G., Zhang, X., . . . Qu, L. J. (2013). Targeted mutagenesis in rice using CRISPR-Cas system. Cell Res, 23(10), 1233-1236. doi:10.1038/cr.2013.123
Miedes, E., Vanholme, R., Boerjan, W., & Molina, A. (2014). The role of the secondary cell wall in plant resistance to pathogens. Front Plant Sci, 5, 358. doi:10.3389/fpls.2014.00358
Miki, B., & McHugh, S. (2004). Selectable marker genes in transgenic plants: applications, alternatives and biosafety. Journal of Biotechnology, 107(3), 193-232. doi:10.1016/j.jbiotec.2003.10.011
Oraby, H. F., & Ramadan, M. F. (2014). Impact of suppressing the caffeic acid O-methyltransferase (COMT) gene on lignin, fiber, and seed oil composition in Brassica napus transgenic plants. European Food Research and Technology, 240(5), 931-938. doi:10.1007/s00217-014-2397-3
Pichon, M., Deswartes, C., Gerentes, D., Guillaumie, S., Lapierre, C., Toppan, A., . . . Goffner, D. (2006). Variation in lignin and cell wall digestibility in caffeic acid O-methyltransferase down-regulated maize half-sib progenies in field experiments. Molecular Breeding, 18(3), 253-261. doi:10.1007/s11032-006-9033-2
Pimentel, D. (2003). Ethanol Fuels: Energy Balance, Economics, and Environmental Impacts Are Negative. Natural Resources Research, 12(2), 127-134. doi:10.1023/a:1024214812527
Piquemal, J., Chamayou, S., Nadaud, I., Beckert, M., Barriere, Y., Mila, I., . . . Pichon, M. (2002). Down-regulation of caffeic acid o-methyltransferase in maize revisited using a transgenic approach. Plant Physiol, 130(4), 1675-1685. doi:10.1104/pp.012237
Renzaho, A. M. N., Kamara, J. K., & Toole, M. (2017). Biofuel production and its impact on food security in low and middle income countries: Implications for the post-2015 sustainable development goals. Renewable and Sustainable Energy Reviews, 78, 503-516. doi:10.1016/j.rser.2017.04.072
Rubinelli, P. M., Chuck, G., Li, X., & Meilan, R. (2013). Constitutive expression of the Corngrass1 microRNA in poplar affects plant architecture and stem lignin content and composition. Biomass and Bioenergy, 54, 312-321. doi:10.1016/j.biombioe.2012.03.001
Sattler, S. E., & Funnell-Harris, D. L. (2013). Modifying lignin to improve bioenergy feedstocks: strengthening the barrier against pathogens? Front Plant Sci, 4, 70. doi:10.3389/fpls.2013.00070
Sattler, S. E., Palmer, N. A., Saballos, A., Greene, A. M., Xin, Z., Sarath, G., . . . Pedersen, J. F. (2012). Identification and Characterization of Four Missense Mutations in Brown midrib 12 (Bmr12), the Caffeic O-Methyltranferase (COMT) of Sorghum. BioEnergy Research, 5(4), 855-865. doi:10.1007/s12155-012-9197-z
Svitashev, S., Schwartz, C., Lenderts, B., Young, J. K., & Mark Cigan, A. (2016). Genome editing in maize directed by CRISPR-Cas9 ribonucleoprotein complexes. Nat Commun, 7, 13274. doi:10.1038/ncomms13274
Svitashev, S., Young, J. K., Schwartz, C., Gao, H., Falco, S. C., & Cigan, A. M. (2015). Targeted Mutagenesis, Precise Gene Editing, and Site-Specific Gene Insertion in Maize Using Cas9 and Guide RNA. Plant Physiol, 169(2), 931-945. doi:10.1104/pp.15.00793
Tang, H. M., Liu, S., Hill-Skinner, S., Wu, W., Reed, D., Yeh, C. T., . . . Schnable, P. S. (2014). The maize brown midrib2 (bm2) gene encodes a methylenetetrahydrofolate reductase that contributes to lignin accumulation. Plant J, 77(3), 380-392. doi:10.1111/tpj.12394
Vad-Nielsen, J., Lin, L., Bolund, L., Nielsen, A. L., & Luo, Y. (2016). Golden Gate Assembly of CRISPR gRNA expression array for simultaneously targeting multiple genes. Cell Mol Life Sci, 73(22), 4315-4325. doi:10.1007/s00018-016-2271-5
Velez-Bermudez, I. C., Salazar-Henao, J. E., Fornale, S., Lopez-Vidriero, I., Franco-Zorrilla, J. M., Grotewold, E., . . . Caparros-Ruiz, D. (2015). A MYB/ZML Complex Regulates Wound-Induced Lignin Genes in Maize. Plant Cell, 27(11), 3245-3259. doi:10.1105/tpc.15.00545
Vignols, F., Rigau, J., Torres, M. A., Capellades, M., & Puigdomènech, P. (1995). The brown midrib3 (bm3) mutation in maize occurs in the gene encoding caffeic acid O-methyltransferase. The Plant Cell, 7(4), 407-416. doi:10.1105/tpc.7.4.407
Wang, G.-F., & Balint-Kurti, P. J. (2016). Maize Homologs of CCoAOMT and HCT, Two Key Enzymes in Lignin Biosynthesis, Form Complexes with the NLR Rp1 Protein to Modulate the Defense Response. Plant Physiology, 171(3), 2166-2177. doi:10.1104/pp.16.00224
Wang, Y., Fan, C., Hu, H., Li, Y., Sun, D., Wang, Y., & Peng, L. (2016). Genetic modification of plant cell walls to enhance biomass yield and biofuel production in bioenergy crops. Biotechnology Advances, 34(5), 997-1017. doi:https://doi.org/10.1016/j.biotechadv.2016.06.001
Yan, J., Yang, X., Shah, T., Sánchez-Villeda, H., Li, J., Warburton, M., . . . Xu, Y. (2010). High-throughput SNP genotyping with the GoldenGate assay in maize. Molecular Breeding, 25(3), 441-451. doi:10.1007/s11032-009-9343-2
Zahradnik, C., Kolm, C., Martzy, R., Mach, R. L., Krska, R., Farnleitner, A. H., & Brunner, K. (2014). Detection of the 35S promoter in transgenic maize via various isothermal amplification techniques: a practical approach. Analytical and Bioanalytical Chemistry, 406(27), 6835-6842. doi:10.1007/s00216-014-7889-2
Zhu, J., Song, N., Sun, S., Yang, W., Zhao, H., Song, W., & Lai, J. (2016). Efficiency and Inheritance of Targeted Mutagenesis in Maize Using CRISPR-Cas9. J Genet Genomics, 43(1), 25-36. doi:10.1016/j.jgg.2015.10.006
|